A Method of Lysate Preparation to Improve the Isolation Efficiency of Protein Partners for Target Proteins Encoded by the Genes of Human Chromosome 18
1Institute of Biomedical Chemistry, , Moscow, 10 Pogodinskaya str., Moscow, 119121 Russian Federation*e-mail: pavel79@inbox.ru
2Institute of Bioorganic Chemistry of the National Academy of Sciences of Belarus, 5, bld. 2 V.F. Kuprevich str., Minsk, 220141 Belarus
Keywords: surface plasmon resonance (SPR); HepG2; lysate preparation; acid treatment and neutralization; protein complexes
DOI:10.18097/BMCRM00090
The aim of this work was to test modifications of the standard protocol for the sample preparation of cell/tissue lysate before performing the affinity isolation of lysate protein partners for the target protein (bait protein) which is covalently immobilized on an inert sorbent (e.g. BrCN-, SH-Sepharose 4B) or a carrier (e.g. paramagnetic nanoparticles). The series of our previous works on applying the approach to direct molecular fishing procedure with combination of affinity chromatography and LC-MS/MS analysis using a number of proteins, encoded by the genes of human chromosome 18, have shown that there are at least two problems affecting the specificity and the effectiveness of this procedure. These include: (i) redundancy of the background proteins in the eluates from an affinity sorbent (carrier) due to isolation of multiprotein complexes “labeled” with a direct protein partner which binds with a bait protein immobilized on the sorbent; (ii) low enrichment of the eluates with appropriate protein partners due to the fact that some direct protein partners in the lysate exist in stable “wild type” complexes with the bait protein itself. This means that latter group of protein partners will not be sufficiently isolated from lysate. Therefore, in order to increase the specificity and efficiency of affinity isolation of protein partners for the bait protein, we modified the standard protocol of lysate preparation and the preliminary step on dissociation of lysate protein complexes was added. Several model experiments for the choice of regeneration solution, assessment of their efficiency in the dissociation of lysate protein complexes as well as the stability and binding capacity of proteins were performed under the control of surface plasmon resonance (SPR) biosensor Biacore 3000 using HepG2 cell lysate. It was shown that acid treatment and incubation of the cell lysate for one min on ice (final lysate dilution 20 times) and subsequent neutralization (pH shift from 2.0 to 7.4) resulted in maximal dissociation of the lysate protein complexes without significant negative effects on the protein-protein interactions tested

|
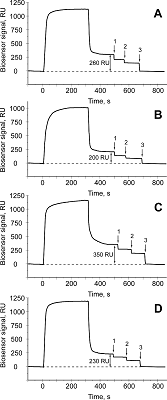
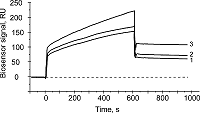
Figure 1.
Testing of regeneration solutions by dissociation efficiency of protein complexes under the SPR control of the Biacore 3000 biosensor ((1) - 2M MgCl2, (2) - 1% CHAPS, (3) - 10 mМ Gly-HCl (pH 2.0)) . Samples of HepG2 cell lysate diluted 10 times with a working buffer were injected on the CM5 chip with immobilized proteins CYB5A (A), RNMT (B), CXXC1 (C), TTR (D) for 5 min at a flow rate of 5 µl/min CYB5A (A), RNMT (B), CXXC1 (C), TTR (D). 1 RU (resonanse unit) corresponds to 1 pg of protein bound to 1 mm2 of the chip surface.
|
|
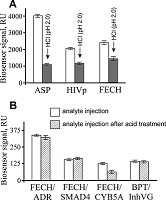
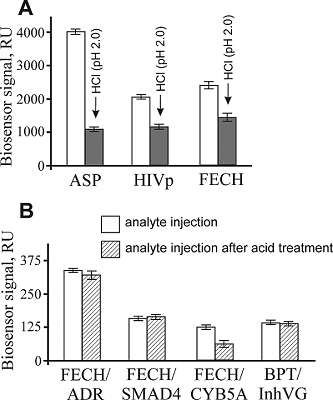
Figure 2.
(A) ─ A decrease of the biosensor signal for three oligomeric proteins immobilized on the optical chip СМ5 after injection of the acid buffer 10 mM Gly-HCl (pH 2.0): L-asparaginase from Erwinia chrysanthemi (ASP), human HIV1 protease (HIVp), human ferrochelatase (FECH).
(B) ─ the biosensor signals of complex formation include by injecting 5 µM different protein analytes [NADPН-dependent adrenodoxin reductase (ADR), mothers against decapentaplegic homolog 4 (SMAD4), microsomal cytochrome b5 (СУВ5А)] via chip surface with immobilized ferrochelatase (FECH), as well as the complex formation between the immobilized bovine pancreatic trypsin (BPT) and peptide inhibitor of VG (InhVG) injected at a concentration of 8 µM. Samples of protein analytes were injected in the absence (control) or after short-term acid treatment (to pH 2.0) followed by subsequent neutralization to pH 7.4. Biosensor signals are shown as the mean values of the biosensor signals ± SD (n = 5). 1 RU (resonance unit) corresponds to 1 pg of protein bound to 1 mm2 of the chip surface. |

|
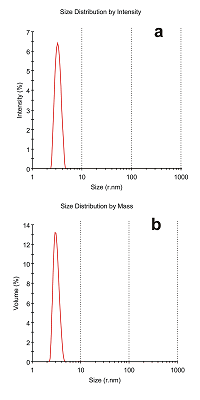
Figure 3.
An example of the particle size distribution analysis by intensity of light scattering (A) and by mass (B) of the protein sample containing ADR protein (Mw 54 kDa) on the ZetaSizer MicroV (Malvern, USA). Additional information: monodisperse peak №1, 3.224 ± 0.432 nm, estimated molecular weight 52.1±7.2, % Intensity 96.8, %Mass 99.3, y-intercept is equal to 0.837 (correlogram).
|
|
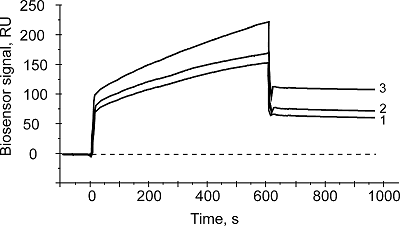
Figure 4.
Typical binding sensograms of protein material from lyzate samples with immobilized on the optical chip cytochrome b5. Next injections are indicated with numerals: (1) - lysate sample (control); (2) - lysate sample treated with 50 mM HCl (followed by subsequent neutralization) with addition of excess of exogenous CYB5A; (3) - lysate sample treated with 50 mM HCl (followed by subsequent neutralization). 1 RU (resonance unit) corresponds to 1 pg of protein bound to 1 mm2 of the chip surface.
|
|
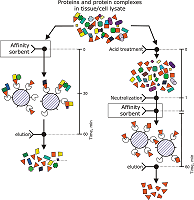
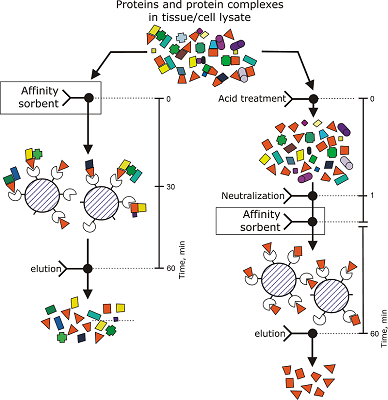
Figure 5.
The schematic representation of a procedure of preparative variant of direct molecular fishing of protein partners for the target protein (covalently immobilized on the inert sorbent) with modified protocol of tissue/cell lysate sample preparation and expected changes in the effectiveness of this procedure.
|
ACKNOWLEDGEMENTS
The reported study was funded by Russian Foundation for Basic Research (RFBR) аccording to the research project №18-04-00071. Authors used the equipment of “Human Proteome” Core Facility (IBMC, Moscow) supported by Ministry of Education and Science of the Russian Federation (unique ID RFMEFI62117X0017).
REFERENCES
- Ershov, P., Mezentsev, Y., Gnedenko, O., Mukha, D., Yantsevich, A., Britikov, V., Kaluzhskiy, L., Yablokov, E., Molnar, A., Ivanov, A., Lisitsa, A., Gilep, A., Usanov, S., Archakov, A. (2012) Protein interactomics based on direct molecular fishing on paramagnetic particles: experimental simulation and SPR validation. Proteomics, 12(22), 3295-3298. DOI
- Ivanov, A.S., Medvedev, A., Ershov, P., Molnar, A., Mezentsev, Y., Yablokov, E., Kaluzhsky, L., Gnedenko, O., Buneeva, O., Haidukevich, I., Sergeev, G., Lushchyk, A., Yantsevich, A., Medvedeva, M., Kozin, S., Popov, I., Novikova, S., Zgoda, V., Gilep, A., Usanov, S., Lisitsa, A., Archakov, A. (2014) Protein interactomics based on direct molecular fishing on paramagnetic particles: Practical realization and further SPR validation. Proteomics, 14(20), 2261-2274. DOI
- Ivanov, A.S., Ershov, P.V., Molnar, A.A., Mezentsev, Yu.V., Kaluzhskiy, L.A., Yablokov, E.O., Florinskaya, A.V., Gnedenko, O.V., Medvedev, A.E., Kozin, S.A., Mitkevich, V.A., Makarov, A.A., Gilep, A.A., Luschik, A.Ya., Gaidukevich, I.V., Usanov S.A. (2016) Direct molecular fishing in molecular partners investigation in protein–protein and protein–peptide interactions. RJBC, 42(1), 14-21. DOI
- Svirid, A.V., Ershov, P.V., Yablokov, E.O., Kaluzhskiy, L.A., Mezentsev, Y.V., Florinskaya, A.V., Sushko, T.A., Strushkevich, N.V., Gilep, A.A., Usanov, S.A., Medvedev, A.E., Ivanov, A.S. (2017) Direct molecular fishing of new protein partners for human thromboxane synthase. Acta Naturae. 9(4), 92-100.
- Florinskaya, A., Ershov, P., Mezentsev, Y., Kaluzhskiy, L., Yablokov, E., Medvedev, A., Ivanov, A. (2018) SPR Biosensors in Direct Molecular Fishing: Implications for Protein Interactomics. Sensors (Basel), 18(5), 1616. DOI
- Gilep, A.A., Guryev. O.L., Usanov, S.A., Estabrook, R.W. (2001) Apo-cytochrome b5 as an indicator of changes in heme accessability: preliminary studies with cytochrome P450 3A4. J. Inorg. Biochem., 87(4), 237-244. DOI
- Usanov, S.A., Graham, S.E., Lepesheva, G.I., Azeva, T.N., Strushkevich, N.V., Gilep, A.A., Estabrook, R.W., Peterson, J.A. (2002) Probing the interaction of bovine cytochrome P450scc (CYP11A1) with adrenodoxin: evaluating site-directed mutations by molecular modeling. Biochemistry, 41(26), 8310–8320. DOI
- Sergeev, G.V., Gilep, A.A. & Usanov S.A. (2014) The role of cytochrome b5 structural domains in interaction with cytochromes P450. Biochemistry (Moscow), 79(5), 406-416. DOI
- Naryzhny, S.N., Maynskova, M.A., Zgoda, V.G., Ronzhina, N.L., Kleyst, O.A., Vakhrushev, I.V., Archakov, A.I. (2016) Virtual-Experimental 2DE Approach in Chromosome-Centric Human Proteome Project. J. Proteome Res., 15(2), 525–530. DOI
- Mezentsev, Yu.V., Molnar, A.A., Gnedenko, O.V., Krasotkina, Yu.V., Sokolov, N.N., Ivanov, A.S. (2006) Oligomerization of L-asparaginase from Erwinia carotovora. Biochem. Moscow Suppl. Ser. B, 1(1), 58-67. DOI
- Mezentsev, Yu.V., Molnar, A.A., Sokolov, N.N., Lisitsina, V.B., Shatskaya, M.A., Ivanov, A.S., Archakov, A.I. (2011) Specificity of Molecular Recognition in Oligomerization of Bacterial L-Asparaginases. Biochem. Moscow Suppl. Ser. B, 5(2), 124-134. DOI
- Ershov, P.V., Gnedenko, O.V., Mol'nar, A.A., Lisitsa, A.V., Ivanov, A.S., Archakov, A.I. (2009) Biosensor analysis of interaction of potential dimerization inhibitors with HIV-1 protease. Biochem. Moscow Suppl. Ser. B, 3(3), 272-288. DOI
- Ershov, P., Mezentsev, Y., Gilep, A., Usanov, S., Buneeva, O., Medvedev, A., Ivanov, A. (2017) Isatin-induced increase in the affinity of human ferrochelatase and adrenodoxin reductase interaction. Protein Sci., 26(12), 2458-2462. DOI
- Fleischmann, G., Fisette, O., Thomas, C., Wieneke, R., Tumulka, F., Schneeweiss, C., Springer, S., Schäfer, L.V., Tampé, R. (2015) Mechanistic Basis for Epitope Proofreading in the Peptide-Loading Complex. J. Immunol., 195(9), 4503-4513. DOI